- Endosymbiote
-
Un endosymbiote est un organisme qui vit à l'intérieur d'une autre cellule ou d'un autre organisme, l'association ainsi formée est une endosymbiose. Pour exemple, la bactérie rhizobium fixatrice d'azote vit dans les nodosités racinaires des légumineuses ou encore l'algue unicellulaire (zooxanthelle) vivant dans les coraux et qui leur permet de construire leur squelette externe ou enfin les bactéries endosymbiotes des insectes qui leur fournissent entre 10 et 15 % de leurs nutriments essentiels, ce qui traduit une large diversité d'endosymbiose. Beaucoup d'endosymbioses sont obligatoires, soit l'hôte soit l'endosymbiote ne peut survivre sans l'autre organisme ou les deux. Mais toutes les endosymbioses ne sont pas obligatoires. Il est également généralement admis que les organites tels les mitochondries et les chloroplastes ont pour origine l'endosymbiose.
La théorie de l'endosymbiose vis-à-vis de la mitochondrie et du chloroplaste
La théorie de l'endosymbiose tente d'expliquer l'origine des organites comme les mitochondries et les chloroplastes chez les cellules eucaryotes. La théorie propose que les chloroplastes et les mitochondries ont évolué à partir de certaines formes bactériennes dont les cellules procaryotes ont été phagocytées. Ces cellules eucaryotes et les bactéries piégées à l'intérieur ont formé des relations d'endosymbiose très serrées et durant très longtemps.
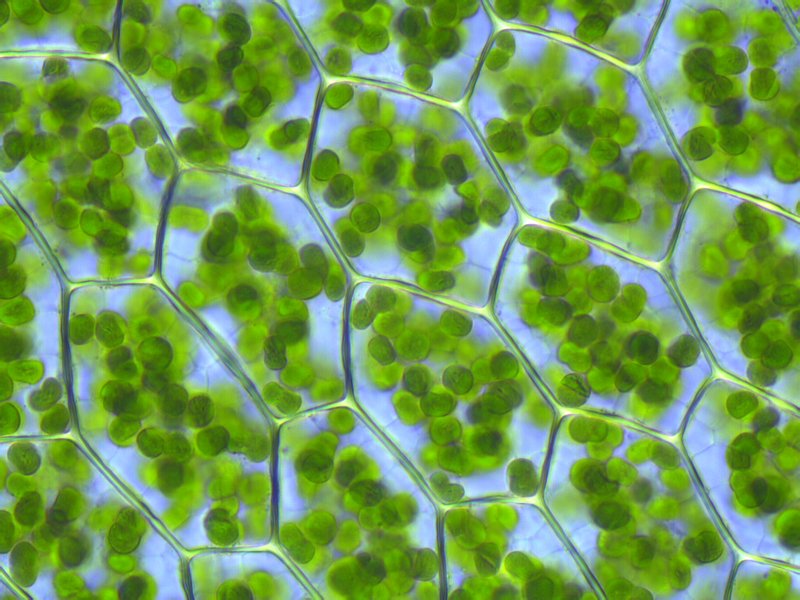
 Chloroplastes dans des cellules végétales de Plagiomnium observées au microscope optique
Chloroplastes dans des cellules végétales de Plagiomnium observées au microscope optique
Suivant la théorie, une cellule anaérobie ingéra une bactérie aérobie mais manqua sa digestion. La bactérie aérobie s'est reproduite dans le cytoplasme (se nourrissant de la composition de ce dernier). Vu que la bactérie avait beaucoup d'énergie, il est probable que certaines avaient de l'ATP (Adénosine Triphosphate) dans leur cytoplasme, chose dont bénéficiait la cellule hôte étant donnée qu'elle était anaérobie stricte. Éventuellement, la bactérie pouvait ne pas pouvoir vivre ailleurs que chez la cellule hôte et ce avant de devenir mitochondrie. Une cellule a dû capturer une cyanobactérie photosynthétique et manqua sa digestion. La bactérie se développa dans le cytoplasme et évolua en chloroplaste primitif. D'autres organites eucaryotes peuvent aussi être le fruit d'une endosymbiose ayant évolué. Des scientifiques pensent que les cils, les flagelles, les centrioles et les microtubules pourraient venir de la symbiose entre une bactérie ciliée et une cellule eucaryote primitive.
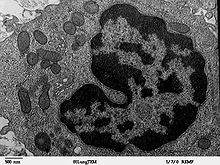 Mitochondries dans un macrophage
Mitochondries dans un macrophageIl y a plusieurs exemples et preuves qui supportent la théorie de l'endosymbiose. Les mitochondries et les chloroplastes contiennent leur propre petit stock d'ADN, qui peut être des vestige du génome de bactéries aérobies indépendantes. La seule preuve la plus convaincante de la descente des organites à partir de bactéries est la fabrication d'arbre phylogénétiques de bactéries concordants entre les mitochondries et des bactéries dite alpha-protéobactéries. Les plastes ont des séquences d'ADN qui indiquent une origine cyanobactérienne. Ce sont des organismes vivant aujourd'hui, appelés êtres vivants intermédiaires qui sont dans une situation similaire d'endosymbiose dans les cellules procaryotes et les bactéries aérobies. Ces êtres vivants intermédiaires montrent que l'évolution proposée par la théorie de l'endosymbiose est possible. Par exemple, l'amibe géante Pelomyxa n'a pas de mitochondrie mais a des bactéries aérobies qui jouent un rôle similaire. Une variété de coraux, de palourdes, d'escargots et une espèce de paramécie ont de manière permanente des algues dans leurs cellules. Beaucoup des endosymbiotes d'insectes ont montré avoir d'anciennes associations avec leurs hôtes, impliquant une inhérence verticale stricte. Ces symbiotes d'insectes ont des modes d'évolution génétique similaires à ceux trouvés dans les vrais organites : réduction du génome, taux rapide d'évolution génétique et des favorisations dans la composition de leurs bases génétiques : favorisant l'adénine et la thymine plutôt que la guanine et la cytosine. D'autres preuves de l'endosymbiose sont les ribosomes procaryotes trouvés au sein de chloroplastes et de mitochondries ainsi que la double membrane qui les entoure. Il était supposé que la membrane interne est la membrane originale du procaryote indépendant, alors que la membrane externe est la membrane de phagocytose. De nos jours, ce point de vue néglige le fait que :
- les cyanobactéries actuelles et les alpha-protéobactéries sont des bactéries à Gram négatif qui sont entourés d'une double membrane
- les membranes externes des organites endosymbiotes (chloroplaste et mitochondrie) sont très similaires à quelques unes de ces bactéries en ce qui concerne leur composition lipidique et protéique[1].
L'accumulation de données biochimiques suggère que la double membrane entourant les chloroplastes et les mitochondries dérive de celles des bactéries ancestrales et la membrane de phagocytose a disparu lors de l'évolution des organites. Des membranes triples ou quadruples sont trouvés chez certaines algues, résultant probablement d'endosymbioses répétés. (Bien que peu d'autres choses ont été retenues de la cellule engloutie). Ces organismes modernes avec des relations d'endosymbiose avec des bactéries aérobies ont vérifié la théorie de l'endosymbiose qui explique l'origine bactérienne des mitochondries et des chloroplastes. Pour les chercheurs en biochimie moléculaire et en évolution biologique il n'est plus question de cette théorie, bien que certains des détails comme les mécanismes de la perte de gènes depuis les organites vers le génome de l'hôte sont toujours étudiés.
Les endosymbioses bactériennes dans les oligochètes marins
Certains oligochètes marins (ex : olavius ou inanidrillus) ont des endosymbiotes extracellulaires obligatoires qui remplissent entièrement le corps de leur hôte. Effectivement, ces vers marins sont nutritionnellement dépendants de leur bactérie symbiote chimiotrophe à défaut d'avoir un système digestif ouvert (pas de tube digestif, pas de bouche, pas de néphridie).
Les endosymbioses bactériennes dans les autres invertébrés marins
Les endosymbiotes bactériens sont aussi représentés dans toutes les classes d'échinodermes. Peu de choses sont connues dans la nature sur les associations (mode d'infection, transmission, besoins métaboliques ...) mais les analyses phylogénétiques montrent que ces symbiotes appartiennent au groupe alpha de la classe des Protéobactéries, relativement de Rhizobium à Thiobacillus. D'autres études montrent que ces bactéries subcuticuliennes pourraient être aussi abondantes sans leurs hôtes et naturellement distribués parmi les Échinodermes en général.
L'endosymbiose dinoflagellés zooxantelles – métazoaire et protozoaires marins
Les endosymbiotes Dinoflagellés du genre Symbiodinium, connus communément sous le nom de zooxanthelles se trouvent dans les coraux, des mollusques (bénitier, Tridacna gigas), des éponges et des foraminifères. Ces endosymbiotes permettent l'étonnante formation des récifs coralliens par la capture de la lumière solaire et donnant à l'hôte l'énergie nécessaire au forçage de la sédimentation carbonatée. Auparavant, ces algues étaient considérés comme une seule espèce, mais phylogénétique moléculaire a mis en évidence les preuves d'une grande diversité de Symbiodinum. Dans certains cas, il y a une spécificité entre l'hôte et la clade du Symbiodinum. Toutefois, le plus souvent il y a une distribution écologique de Symbiodinum, le symbiote peut commuter son hôte avec une apparente facilité. Quand les récifs deviennent environnementalement stressés, cette distribution des symbiotes est reliée à la tendance observée de blanchissement des coraux. Donc la distribution de Symbiodinum sur les récifs de coraux et son rôle dans le blanchissement des coraux présente un des plus importants complexes et un problème courant intéressant dans l'écologie des récifs.
L'endosymbiose chez les protozoaires
Mixotricha paradoxa est un protiste qui n'a pas de mitochondrie, en revanche, une bactérie sphérique vit en son sein et compense les fonctions manquantes des mitochondries. Mixotricha a aussi trois autres espèces de symbiotes qui vivent à la surface de sa cellule. Paramecium bursaria, une espèce de cilié entretient une symbiose mutualiste avec une algue verte appelée zoochlorelle. Cette algue vit dans le cytoplasme du cilié.
L'endosymbiose bactérienne obligatoire chez les insectes
Les scientifiques classifient les endosymbiotes d'insectes en deux catégories, primaire et secondaire. Les endosymbiotes primaires (parfois référés comme P-endosymbiotes) ont été associés avec leur insecte hôte depuis des millions d'années (de dix à plusieurs centaines de millions d'années dans certains cas). Ils ont une forme d'association obligatoire et ils affichent une co-spéciation avec leur hôte. Les endosymbiotes secondaires exposent une association développée plus récemment, qui est parfois horizontalement transférée entre les hôtes, vivant dans l'hémolymphe des insectes (les bactériocytes non spécialisés), et ils ne sont pas obligatoires.
 pucerons
puceronsParmi les endosymbiotes primaires des insectes, le plus étudiés sont les pucerons du pois (Acyrthosiphon pisum) endosymbiote Buchnera sp[2]. La mouche tsétsé Glossina morsitans morsitans et son endosymbiote Wiggleworthia glossinidia brevipalpis et le protiste endosymbiote des termites. Comme avec l'endosymbiose chez les autres insectes, la symbiose est obligatoire c'est-à-dire que ni la bactérie ni l'insecte ne peut vivre sans son hôte (resp. son symbiote). Les scientifiques ont été incapables de cultiver la bactérie en laboratoire dans des conditions externes à l'insecte. Avec des rations spéciales, l'insecte peut survivre mais est en mauvaise santé et au mieux ne survit que quelques générations.
Dans certains groupes d'insectes, ces endosymbiotes vivent dans des cellules d'insectes spécialisés appelées bactériocytes (aussi appelées mycétocytes) et sont maternellement transmis i.e. La mère transmet son endosymbionte à sa progéniture. Dans certains cas, la bactérie est transmise dans l'œuf, comme Buchnera ; dans d'autres cas comme Wigglesworthia, ils sont transmis par le lait qui permet le développement de l'embryon? Chez les termites, l'endosymbiote réside dans les intestins et sont transmis par trophallaxie avec les autres membres de la colonie. Les endosymbiotes primaires sont là pour aider l'hôte en lui permettant d'avoir des nutriments que l'hôte ne pourrait obtenir seul, en catabolisant la nourriture inassimilable de l'insecte. Par exemple, le rôle élémentaire de Buchnera des de synthétiser les acides aminés essentiels que le puceron ne peut naturellement obtenir avec sa nourriture. De la même manière, le rôle essentiel de Wigglesworthia est probablement de synthétiser des vitamines dont la mouche tsétsé ne dispose pas dans le sang avec lequel elle se nourrit. Chez les termites, le protiste endosymbiote joue un rôle majeur dans la digestion du matériel ligneux et cellulosique qui constitue la nourriture des termites. Les bactéries bénéficient d'une exposition réduite aux prédateurs, la fourniture de suffisamment de nutriments et de la stabilité de l'environnement immédiat qui est le corps de l'hôte. Le séquençage du génome révèle que les bactéries endosymbiotes obligatoires des insectes ont le plus petit des génomes bactériens connus et ont perdu de nombreux gènes qui sont habituellement trouvés dans de nombreuses bactéries. Plusieurs théories ont été mises en avant pour expliquer la perte de ces gènes. Probablement certains de ces gènes ne sont pas indispensables dans l'environnement de la cellule dans l'insecte hôte. Une théorie complémentaire suggère que le petit nombre de bactéries dans chaque insecte diminue l'efficacité de la sélection naturelle en « purgeant » les mutations délétères et les petites mutations de la population, résultant dans la perte de gènes pendant plusieurs millions d'années. Les recherches dans une phylogénie parallèle entre les insectes et leurs symbiotes ont déduit que l'endosymbiote primaire est transféré seulement verticalement (i.e. par la mère), et pas horizontalement (i.e. en s'échappant de l'hôte et entrant dans un autre). Les attaques obligatoires des bactéries endosymbiotes peuvent être une voie pour contrôler leurs insectes hôtes dont beaucoup sont des ravageurs ou des porteurs de maladie humaine. Par exemple, les pucerons sont des ravageurs des cultures et la mouche tsétsé transmet l'organisme Trypanosoma gambiense qui cause la maladie du sommeil. D'autres motivations de ces études sont de comprendre de la symbiose et de comprendre comment des bactéries avec des lacunes génomiques peuvent vivre, ainsi que d'améliorer notre connaissance de la génétique et de la biologie moléculaire. Moins de choses sont connues sur les endosymbiotes secondaires. Le puceron du pois est connu pour contenir au moins trois endosymbiotes secondaires Hamiltonella defensa, Regiella insecticola, et Serratia symbiotica. H. defensa aide à la défense de l'insecte contre les parasitoïdes. Sodalis glossidinus est un endosymbiote secondaire de la mouche tsétsé qui vit à l'extérieur et à l'intérieur des cellules dans différents tissus de son hôte, incluant les intestins et l'hémolymphe. Les études phylogénétiques n'indiquent pas de corrélation entre l'évolution de Sodalis et la mouche tsétsé, contrairement à Wrigglesworthia, même si Sodalis a été cultivé in vitro[3] Unlike tsetse's P-symbiont Wigglesworthia, though, Sodalis has been cultured in vitro[4].
Les endosymbiotes viraux, rétrovirus endogènes

Pendant la gestation des mammifères vivipares, les endorétrovirus sont activés et sont produits en grande quantité pendant l'implantation de l'embryon. D'un côté, ils activent les immunodépresseurs et protègent l'embryon du système immunitaire de la mère et d'un autre côté, la fusion des protéines virales provoque la formation du syncytium placentaire en vue de limiter les échanges et les migrations de cellules entre l'embryon en développement et le corps de la mère, un épithélium ne le fait pas car certaines cellules du sang sont spécialisées pour s'insérer entre les cellules épithéliales. L'endorétrovirus est un virus similaire à celui du HIV. L'action immunodépressive était l'activité initiale du virus, comme le HIV. La fusion des protéines était le moyen de répandre l'infection aux autres cellules simplement en répandant le virus. Il est pensé que les ancêtres des animaux vivipares actuels ont évolué après une infection accidentelle avec un ancêtre du virus actuel, ce qui a permis au fœtus de survivre au système immunitaire de sa mère[5]. Le projet génome humain découvre quelques milliers d'entérorétrovirus qui sont organisés en 24 familles[6].
Bibliographie
Les endosymbioses bactériennes dans les oligochètes marins
Endosymbiotic sulphate-reducing and sulphide-oxidizing bacteria in an oligochaete worm. Dubilier N., Mülders C.,Ferdelman T., De Beer D.,Pernthaler A.,Klein M., Wagner M., Erseus C., Thiermann F., Krieger J., Giere O & Amann R. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=11357130
Endosymbiose chez les Échinodermes
Subcuticular bacteria from the brittle star Ophiactis balli (Echinodermata: Ophiuroidea) represent a new lineage of extracellular marine symbionts in the alpha subdivision of the class Proteobacteria. Burnett, W J and J D McKenzie http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=168468&rendertype=abstract
Endosymbiose chez les Échinodermes marins et protozoaires Excellent review paper covering the role of Symbiodinium in reef ecology and the current state of research: FLEXIBILITY AND SPECIFICITY IN CORAL-ALGAL SYMBIOSIS: Diversity, Ecology, and Biogeography of Symbiodinium. Andrew C. Baker, Annual Review of Ecology, Evolution, and Systematics 2003 34, 661-689
Endosymbiose chez les insectes
PLOS Biology Primer- Endosymbiosis: lessons in conflict resolution http://www.plosbiology.org/plosonline/?request=get-document&doi=10.1371/journal.pbio.0020068 A general review of bacterial endosymbionts in insects. P. Baumann, N. A. Moran and L. Baumann, Bacteriocyte-associated endosymbionts of insects in M. Dworkin, ed., The prokaryotes, Springer, New York, 2000. http://link.springer.de/link/service/books/10125/ An excellent review of insect endosymbionts that focuses on genetic issues. Jennifer J. Wernegreen (2002), Genome evolution in bacterial endosymbionts of insects, Nature Reviews Genetics, 3, pp. 850-861. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=12415315&dopt=Abstract A review article on aphids and their bacterial endosymbionts. A. E. Douglas (1998), Nutritional interactions in insect-microbial symbioses: Aphids and Their Symbiotic Bacteria Buchnera, Annual Reviews of Entomology, 43, pp. 17-37. Describes possible methods to control the human pathogen causing African sleeping sickness, which is transmitted by tsetse flies. Focuses on methods using the primary and secondary endosymbionts of the tsetse fly. Serap Aksoy, Ian Maudlin, Colin Dale, Alan S. Robinsonand and Scott L. O'Neill (2001), Prospects for control of African trypanosomiasis by tsetse vector, TRENDS in Parasitology, 17 (1), pp. 29-35. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=11137738&dopt=Abstract Announces and analyzes the full genome sequence of Buchnera sp. APS, the endosymbiont of the pea aphid, and the first endosymbiont to have its genome sequenced. S. Shigenobu, H. Watanabe, M. Hattori, Y. Sakaki and H. Ishikawa (2000), Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. APS, Nature, 407, pp. 81-86. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=10993077&dopt=Abstract An article that presents for the first time a theory on how obligate endosymbionts may have their genomes degraded, in a freely-available journal. Nancy A. Moran (1996), Accelerated evolution and Muller's ratchet in endosymbiotic bacteria, Proceedings of the National Academy of Sciences of the USA, 93, pp. 2873-2878. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=8610134&dopt=Abstract
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « endosynbiont » (voir la liste des auteurs)
- K Inoue, « The chloroplast outer envelope membrane: the edge of light and excitement », dans Journal of Integrative Plant Biology, vol. 49, 2007, p. 1100–1111
- A E Douglas, « Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic bacteria Buchnera », dans Annual Review of Entomology, vol. 43, 1998, p. 17–38 [lien DOI]
- Aksoy, S., Pourhosseini, A. & Chow, A. 1995. Mycetome endosymbionts of tsetse flies constitute a distinct lineage related to Enterobacteriaceae. Insect Mol Biol. 4, 15-22.
- Welburn, S.C., Maudlin, I. & Ellis, D.S. 1987. In vitro cultivation of rickettsia-like-organisms from Glossina spp. Ann. Trop. Med. Parasitol. 81, 331-335.
- [1] The Viruses That Make Us: A Role For Endogenous Retrovirus In The Evolution Of Placental Species (by Luis P. Villarreal)
- [2] Persisting Viruses Could Play Role in Driving Host Evolution
Voir aussi


Wikimedia Foundation. 2010.